| Chromatography of Plant Pigments |  |
Introduction
Chromatography is a way of separating a mixture using differences in the abilities of the components to move through a material. All chromatography involves two phases – a stationary phase and a mobile phase. The movement of the mobile phase through the stationary phase allows separation to take place. Because the components of a mixture move at different rates, they eventually separate.
Paper chromatography is a common way to separate various components of a mixture. The components of the mixture separate because different substances are selectively absorbed by paper due to differences in polarity. A solution can be separated by allowing it to flow along a stationary substance. Water or some other solvent is used as the mobile phase. The solvent moves upward along the paper because of capillary action. As it reaches the spot, the mixture dissolves in the solvent. For instance, the pigments in an ink solution can be separated by passing the ink through a piece of paper. The pigments respond differently to the paper. The differences in the migration rates result in differences in the distances the separated components travel, some pigments are held back while other moves ahead. Eventually, a pattern of colors results that shows the separated pigments.
Hypothesis
Paper can be used to separate mixed chemicals.
Materials
The materials used for this lab are paper, pencil, scissors, eraser, filter paper, test tube, cork, paper clip, metric ruler, black felt-tip pen, and a calculator.
Methods
The first step to this experiment was to bend a paper clip so that it is straight with a hook at one end. Push the straight end of the paper clip into the bottom of a cork stopper. Next, hang a thin strip of filter paper on the hooked end of the paper clip. Insert the paper strip into the test tube so it does not touch the sides, but almost the bottom of the test tube. Next, remove the paper strip from the test tube and draw a solid 5 mm wide band about 25 mm from the bottom of the paper, using a black felt tip pen. Use a pencil to draw a line across the top of the paper strip 10 cm from the top.
Pour about 2 mL of water into the test tube with the bottom of the paper in the water and the black band above the water. Observe what happens as the liquid travels up the paper. Record the changes you see. When the solvent has reached the pencil line, remove the paper from the test tube. Let the paper dry on the desk. With a metric ruler, measure the distances form the starting point to the top edge of each color. Record the data in a data table. Calculate a ration for each color by dividing the distance the color traveled by the distance the solvent traveled.
Results
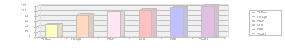
The results of the experiment are shown in a chart and a graph.
Distance color traveled and Rf value.
| Color of ink (list in order | Distance traveled by each color (mm) | Distance solvent traveled (mm) | Ration traveled = Distance color moved /Distance water moved |
| Yellow | 50 | 120 | 5/12 |
| Orange | 85 | 120 | 17/24 |
| Pink | 100 | 120 | 5/6 |
| Red | 105 | 120 | 7/8 |
| Blue | 115 | 120 | 23/24 |
| Violet | 120 | 120 | 1 |
Questions
1. How many colors separated from the black ink? Six colors separated from the black ink: yellow, orange, pink, red, blue, violet.
2. What served as the solvent for the ink? Water served as the solvent because it is the universal solvent.
3. As the solvent travel up the paper, what color appeared first? Orange appeared first as the solvent traveled up the paper.
4. List the colors in order from top to bottom that separated from the black ink? The colors that separated from top to bottom: violet, blue, red, pink, orange, and yellow.
5. In millimeters, how far did the solvent travel. The solvent traveled 120mm.
6. From your results, what can you conclude is true about black ink. That black ink is a combination of several colors and that can be separated by water.
7. Why did the inks separate? The ink separated because each pigment has its own characteristics and molecular structure.
8. Why did some inks move a greater distance? Different pigments were absorbed at different rates.
Error analysis
There could be an error by the way the ink was distributed on the paper or by the amount of water put in the test tube.
Conclusion
The hypothesis was correct. This experiment showed the way black ink could be separated. Black ink is made from a various colors— yellow, orange, pink, red, blue, and violet. The colors separate because of the differences in their molecular characteristics, their solubility in water and their rate of absorption by the paper.