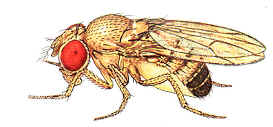
The mega-nosed fly (Moegistorhynchus longirostris) of southern Africa, like its literary counterpart, Pinocchio, has a bizarre appearance that reveals an underlying truth. Its proboscis, which looks like a nose but is actually the longest mouthpart of any known fly, protrudes as much as four inches from its head–five times the length of its bee-size body. In flight the ungainly appendage dangles between the insect’s legs and trails far behind its body.
To an airborne fly, an elongated proboscis might seem a severe handicap (imagine walking down the street with a twenty-seven-foot straw dangling from your mouth). Apparently, though, the handicap can be well worth its aerodynamic cost. The outlandish proboscis gives the mega nosed fly access to nectar pools in long, deep flowers that are simply out of reach to insects with shorter mouthparts.
But that poses a conundrum: why would natural selection favor such a deep tube in a flower? After all, nectar itself has evolved because it attracts animals that carry pollen, the sperm of the floral world, from one plant to another. And since pollinators perform such an essential service for the flower, shouldn’t evolution have favored floral geometries that make nectar readily accessible to the pollinators?
Yet the story of the long proboscis of the mega-nosed fly and the long, deep tubes of the flowers on which it feeds is not quite so straightforward. There are subtle advantages, it turns out, to making nectar accessible to only a few pollinators, and nature factors those advantages into the evolutionary equation as well. In fact, the evolution of those two kinds of organisms, pollinator and pollinated, presents an outstanding example of an important evolutionary phenomenon known as coevolution. Coevolution can explain the emergence of bizarre or unusual anatomies when no simple evolutionary response to natural selection is really adequate. It can help conservationists identify species that could be vital in maintaining a given habitat. And it can help naturalists investigating novel plants predict what kinds of animals might pollinate their flowers.
The coevolution of the mega nosed fly and the plants it pollinates is a tale of extreme specialization. Each species has adapted to changes in the other in ways that have left each of them, to some degree, reliant on the other. The idea that a plant species might become dependent for pollination on a single species of animal goes back to the writings of Charles Darwin. For example, Darwin noted, the flower spur of the Malagasy orchid (Angraecum sesquipedale) contains a pool of nectar that is almost a foot inside the opening of the flower. (A flower spur is a hollow, hornlike extension of a flower that holds nectar in its base.) In pondering the evolutionary significance of those unusual flowers, Darwin predicted that the orchid must be adapted to a moth pollinator with a long proboscis.
Critical to Darwin’s prediction was his suspicion that pollination could take place only if the depth of a plant’s flowers matched or exceeded the length of a pollinator’s tongue. Only then would the body of the pollinator be pressed firmly enough against the reproductive parts of the flower to transfer pollen effectively as the pollinator fed. Thus, as ever deeper flowers evolved through enhanced reproductive success, moths with ever longer proboscises would also, preferentially, live long enough to reproduce, because they would most readily reach the available supplies of nourishing nectar. Longer proboscises would lead yet again to selection for deeper flower tubes.
The result would be the reciprocal evolution of flowers and pollinator mouthparts. That coevolutionary process would cease only when the disadvantages of an exaggerated trait balanced or outweighed its benefits. Given enough time, the process might even produce new species: an insect the specializes in feeding on nectar from deep flowers, and a deep-flowered plant specialized for being pollinated by insects with long mouthparts.
In the early twentieth century it seemed that Darwin’s prediction had been borne out. A giant hawk moth from Madagascar, Xanthopan morganii praedicta, was captured, with a proboscis that measured more than nine inches long. Although no one has actually seen the insect feeding on the flower, the discovery is still remarkable, and strongly suggestive of the coevolution of the orchid and moth. Other insects that have relationships with highly specific plants, such as the mega nosed fly and other, related long-nosed fly species of southern Africa, provide even better evidence of the reciprocal links between planes and their pollinators.
Darwin would have been amazed that some flies in southern Africa have longer tongues than most hawk moths do. After all, the flies’ bodies are several times smaller than the hawk moths’ are. Flies are described as long-nosed if their mouthparts are longer than three quarters of an inch. By that criterion, more than a dozen long-nosed fly species are native to southern Africa. They belong to two families. The nemestrinids, or tangle-veined flies (which include the mega-nosed fly), feed solely on nectar, whereas the tabanids, or horseflies, feed mostly on nectar, though female tabanids have separate mouthparts to suck blood for their developing eggs.
Like all other long-nosed flies, the mega nosed fly is the sole pollinator to a group of unrelated plant species; such a group is known as a guild. The plant guild of the mega nosed fly includes species from a wide variety of plant families, including geraniums, irises, orchids, and violets.
Even though guild members may be only distantly related, all of them have roughly the same characteristics. For example, plants in the long-nosed fly guild all have long, straight floral tubes or spurs; brightly colored flowers that are open during the day; and no scent. The defining traits of a guild together form what botanists call a pollination syndrome. For example, bird-pollinated flowers are typically large, red, and unscented, whereas moth-pollinated flowers are more likely to be long, narrow, white, and scented in the evening.
The most important trait in the pollination syndrome of the long-nosed fly (and indeed, in all pollination syndromes of long-nosed insects) is a deep, tubular flower or floral spur. One of us (Johnson) and Kim E. Steiner of the Compton Herbarium in Claremont, South Africa, studied the orchid Disadraconis, a southern African plant with a deep, tubular floral spur. The two investigators artificially shortened the spurs of some orchids in a habitat where the only pollinators present were long-nosed flies. The plants whose spurs remained long got more pollen, and were more likely to produce fruits, than the ones whose spurs were shortened.
Yet short floral spurs are not necessarily a reproductive disadvantage. Shorter spurs would make it possible for a wider range of pollinators to access the nectar, if various potential pollinators are present. Instead, longer spurs only seem to be an advantage when long-tongued insects are the sole pollinators. Johnson and Steiner found that differences in spur length among populations cannot be blamed on differences in moisture or temperature, thus reinforcing their conclusion that spur length was an adaptation to the local distributions of long-tongued flies.
Not only does spur length correlate statistically with pollinator traits, but a direct causal connection can be demonstrated. Johnson and Ronny Alexandersson, a botanist at Uppsala University in Sweden, studied South African Gladiolus flowers pollinated by long-tongued hawk moths. When the hawk moth proboscises were long compared to the length of the flower tube, the hawk moths did not efficiently pick up pollen, and the flowers did not reproduce well. When the hawk moth proboscises were relatively short, pollen was more readily transferred, and the plants were more likely to be fertilized and bear fruit. Thus the length of the pollinator’s proboscis exerts a strong pressure on the reproductive success of the flowers.
Those studies and others suggest that what Darwin predicted of the Malagasy orchid is a rather general phenomenon: hawk moths and long-nosed flies coevolved with their plant partners. As floral tubes became longer, so did the pollinators’ proboscises, and those led, in turn, to even longer flowers. As the lengths of the flower tube and the insect proboscis converge, a remarkable degree of specialization develops. The plants come to rely for pollination on the few insect species that can reach their flowers’ nectar supplies.
There are advantages for the specialists on both sides of this relationship. The long-nosed flies obviously get privileged access to pools of nectar. And the plants pollinated by long-nosed flies benefit from a near-exclusive pollen courier service–or at least one that minimizes the risk of delivery to the wrong address. But specializing can also be a risky strategy for the plants if the pollinators are less interested in fidelity than the plants are. Long-nosed flies could not survive on the nectar they could get by visiting just one plant species; the flies must visit several plant species to gather the energy they need. Johnson and Steiner observed mega nosed flies visiting at least four species with deep flowers.
Such promiscuous behavior could be detrimental to the plants. A fly might end up carrying pollen from one species to a different species in the guild, thereby wasting the pollen. Worse, the foreign pollen could end up clogging the stigmata, the female reproductive structures, of the receiving flowers, preventing them from getting the “right” pollen. But the stigmata of plants in the guild of the mega nosed fly do not clog, because among those plants yet another clever adaptation to specialized pollination has evolved. Each plant species arranges its anthers, the male reproductive structures, in a characteristic position. That way, the pollen from each species sticks to the pollinator’s body in a distinct but consistent, plant-specific location. The fly becomes an even more efficient courier, carrying pollen from various plant species simultaneously, say, on its head, legs, and thorax.
The risks of specialization are not confined to the flowers. Just as the flies are unfaithful partners, some flowers are dishonest about signaling a nectar reward. The orchid D. draconis, for instance, is not the mutualistic partner it seems. The flower attracts the mega-nosed fly because it looks like other members of the fly’s guild. But, whereas the fly carries the orchid’s pollen, the orchid offers no nectar in return.
The risk of falling for such a trick seems a small price for the flies to pay for the benefits of specialization. But specialization also carries a much graver risk–in fact the ultimate risk–for both members of the partnership because the disappearance of either partner is likely to doom the other one, as well. Some plant species have mechanisms, such as vegetative reproduction or self-pollination, that may help sustain their populations in the short run. But in the long run, without their pollinators, the species will slowly and irrevocably decline. Pollinating insects may be more flexible in some cases, but are still vulnerable if a key food source disappears.
Unfortunately, in southern Africa that is just what is happening to many plants and their long-nosed fly partners. Often not even closely related insect species can help in pollination. For affected plants, the loss of a single fly species means extinction. And examples of that gloomy cascade have already been observed. Peter Goldblatt of the Missouri Botanical Garden in St. Louis and John C. Manning of the Compton Herbarium have ‘reported that many populations of long-nosed flies are threatened by the loss of their wetland breeding habitat, and also, possibly, by the loss of other insects they parasitize during their larval stages. In some habitats, flowers in the long-nosed fly guild already produce no seeds, because their pollinator is locally extinct.
Naturalists have accepted the concepts of guilds and pollinator syndromes for many years, and predicting which pollinators regularly visit which plants has become something of a cottage industry. But just how common is pollinator specialization in southern Africa? Promiscuity could turn out to be a more successful–and more widespread–strategy than specialization, even among plants that seem to fit into identifiable guilds.
In recent years ecologists have discovered that just because plants and insects appear to form a pollination guild does not guarantee they never venture outside it. For example, ecologists have noted that in years when hummingbird populations are low, flowers ordinarily pollinated by hummingbirds can fill up with nectar and become pollinated effectively by bees. Likewise, bees once thought to specialize in only one or two plant species turn out to forage on a variety of plants.
The take-home lesson has been that the syndrome concept is no substitute for careful field observation. Some investigators even think that the concept has caused botanists to overlook generalists. In the Northern Hemisphere, for instance, studies suggest that generalization is the norm, not the exception. Johnson and Steiner recently completed a study showing that members of the orchid and asclepiad families in the Northern Hemisphere tend to rely on between three and five pollinators each. In contrast, plants from the same families in the Southern Hemisphere rely on just one pollinator each.
So why might generalization be more common in the Northern Hemisphere than it is in the Southern Hemisphere? Perhaps the reason is that social bees, which are largely opportunistic, dominate pollinator faunas in northern regions. In the Southern Hemisphere, by contrast, social bees are mostly absent, replaced instead by more specialized pollinators such as the long-nosed flies and hawk moths.
But that is just a broad generalization itself. More data on the geographic distribution of pollinator specialization needs to be gathered, particularly in tropical countries. The data is vital, not only to advance the specialization debate, but also to protect as many of these unique species and relations as possible, lest they disappear forever.